生物竞赛的同学借了一本书《动物行为学(第二版)》,在里面找到了一些对于trans可能有点用的内容,特此摘录。
这是动物行为学,虽然人属于哺乳动物有一定参考性,但是不特指人哦。
不知道放在哪里好,就先放在脱水菜吧。
关于这些内容的解析与要点我会放到第二帖。
注意,对于一些和Trans相关度较低的内容,锦心做了删去。文字由识图生成,可能有一定和原文不符合,欢迎指出。
如果你在生物竞赛或者对其它内容很感兴趣,欢迎购买这本书。
本文仅限于学习交流只用,侵权删除。如果有下载请在24小时内删除。
下面是正文。
二、脊椎动物的内分泌系统(P127)
脊推动物的内分泌系统有什么特点?它们与无脊椎动物的内分泌系统又有什么不同呢?在进化过程中很多变化都是发生在动物的内分泌系统。特别是脊椎动物的内分泌系统已经进化产生了两个主要成分:①脑下垂体,它位于脑腹面的下丘脑附近并与几个中枢神经系统的结构密切相连。脑下垂体和下丘脑是紧密相连的,共同形成了神经和内分泌系统之间的重要桥梁。这个桥梁对于这两个控制系统之间的整合是极为重要的,其中存在着两个连接:垂体后叶主要是由来自下丘脑的神经元组成的,而垂体前叶则靠下丘脑-垂体门静脉系统与下丘脑相连。脑下垂体产生促激素影响其他内分泌腺,同时也产生直接起作用的激素。②其他内分泌腺体,包括甲状腺、松果体、肾上腺、胰腺和生殖腺,这些腺体位于身体的各处。由垂体各个部位所释放的促激素就是肽(peptides),肽的基本结构是由氨基酸链组成的。而来自肾上腺、精巢、卵巢和胎盘的激素属于类固醇激素(steroid hormones),这类激素都有碳环结构并生有各种附加的侧链。
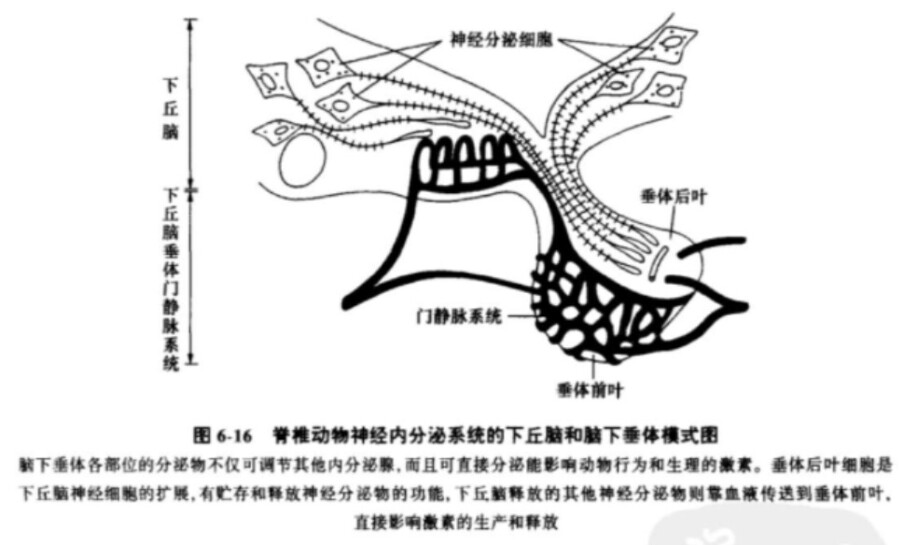
脑下垂体各部位的分泌物,不仅可调节其他内分泌腺,而且可直接分泌能影响动物行为和生现的激素。垂体后叶细胞是下丘脑神经细胞的扩展,有贮存和释放神经分泌物的功能,下丘脑释放的其他神经分泌物则靠血液传送到垂体前叶,直接影响激素的生产和释放
1.内分泌物及其对生理和行为的影响
脊椎动物的主要内分泌腺都有哪些种类以及它们是如何影响动物行为的呢?现在已知脑下垂体的几种分泌物对脊椎动物的行为和生理机制起着调控作用(表6-2)。催产素(oxytocin)和升压素(vasopressin)是下丘脑神经元生产的并贮存在垂体后叶神经终端,然后再作为神经分泌物被释放到血液中。催产素的作用是刺激子宫收缩,有助于交配后精子在雌性生殖道内的移动,而且也有助于分娩期间驱动胎儿。催产素也能刺激乳汁从乳腺中泌出。升压素则影响着肾脏的生理功能并可改变尿液浓度,从而有助于调节水分平衡。例如,高浓度尿液的排泄和身体水分的贮存是很多荒漠哺乳动物的生理和行为适应,如骆驼、更格芦鼠和沙鼠等。
表6-2 脊椎动物激素的种类、来源及其对动物生理和行为的影响
| 激素名称 | 来源 | 对生理和行为的影响 |
|---|---|---|
| 褪黑激素 | 松果体 | 调节年生殖周期 |
| 催产素、升压素 | 垂体后叶 | 排奶,催产,水分平衡 |
| 促黄体素(LH) | 垂体前叶 | 形成黄体;分泌孕酮;分泌雄激素 |
| 促卵泡激素(FSH) | 垂体前叶 | 卵泡发育;排卵(与LH和雌激素一起) |
| 促乳素 | 垂体前叶 | 分泌乳汁;鸟类双亲抚育行为 |
| 促肾上腺皮质激素(ACTH) | 垂体前叶 | 分泌肾上腺类固醇 |
| 促黑激素(MSH) | 垂体中叶 | 变色 |
| 类固醇 | 肾上腺皮质 | 水分平衡;代谢;电解质平衡 |
| 肾上腺素,去甲肾上腺素 | 肾上腺髓质 | 血糖水平;压力反应 |
| 雄性激素 | 精巢 | 精巢发育;精子发生;第二性征 |
| 雌性激素 | 卵巢和胎盘 | 子宫发育;乳腺发育 |
| 孕激素 | 卵巢和胎盘 | 维持妊娠 |
可间接影响动物行为的四种激素是由垂体前叶分泌的,其中三种是可影响其他内分泌腺的促激素。在雌性动物中,促卵泡激素(FSH)和促黄体素(LH)可影响卵巢中卵的成熟周期、性接受力和妊娠。在雄性动物中,FSH和LH可控制精子的生成和雄性激素的分泌。第三种促激素是促肾上腺皮质激素( adrenocorticotrophic hormone,ACTH ) ,它可影响肾上腺皮质类固醇醇激素的生产和分泌。存在于鸟类和哺乳动物的一种 脑垂体激素促乳索 (prolactin)对母爱行为是很重要的,它能影响哺乳动物乳汁的生产和鸟类嗉囊乳的累积。在某些两栖动物中也曾发现过促乳素,它的功能可能是促使迁往有水的地方进行生殖。脑内的松果体可分泌多种激素包括吲哚胺( indoleamine)、蛋白质和多肽( polypeptides)。对动物行为研究来说,其中最重要的是褪黑激素,它的作用是调节哺乳动物的生殖和年生殖活动格局。
生殖腺是由来自垂体的营养分泌物所激活的,使精巢分泌雄激素,卵巢生产雌激素和孕激素。在妊娠期间胎盘所分泌的孕激素对维持妊娠起着关键作用。这些激素不仅影响着动物的生殖、母爱行为和群聚,而且也决定着动物的某些起着通讯信号作用的第二性征。肾上腺激素与保持水分平衡、维持新陈代谢和电解质平衡有着密切关系。来自肾上腺髓质的肾上腺素( adrenaline )和去甲肾上腺素( noradrenaline )在突发的压力反应中起着重要作用。
2.内分泌腺之间的相互作用
很多内分泌腺的分泌活动都具有反馈性质,即具有反馈环( feedback loops),如图所示。
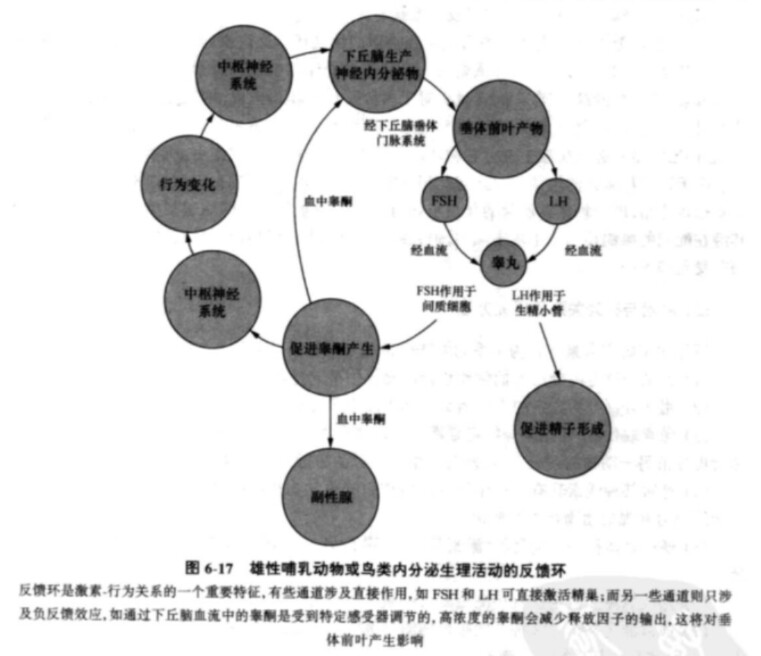
脑垂体分泌的FSH和LH是受释放因子调控的,后者来自于下丘脑并流经下丘脑垂体门静脉系统。此后,促激素FSH和LH被送入血液输送到精巢并在那里激活生精小管的精子发生过程,使间质细胞产生和释放睾酮。反过来睾酮又会进入血液传送到其他部位,包括副性腺和下丘脑。特化的下丘脑感觉细胞是构成身体内稳定机制的一部分并不断监测着血液中各种化学物质的浓度,其中包括睾酮及其代谢产物。因此当一个动物受到阉割时,它的睾酮含量就会下降,但同时FSH的浓度会增加。在这种情况下如果动物的行为发生了变化,那我们应当把这种变化归之于睾酮浓度的下降还是FSH浓度的增加呢?这点也正是我们对影响行为的激素进行研究的难点之一。
处于循环状态的率酮浓度影响着下丘脑往脑垂体分部释放因子,这是一种负反馈关系。当血液中睾酮浓度增加时,下丘脑释放因子的分泌就会减少。相反,当血液中睾酮浓度下降时,经由下丘脑-垂体门静脉系统分泌到垂体前叶的释放因子就会增加并会引起FSH和LH输出的增加。在FSH、LH和雌性生殖激素如雌激素(estrogen)和孕激素( progesterone)之间也存在着反馈环,这种反馈环可用雌性哺乳动物的动情周期( estrous cycle )加以说明。在动情周期开始时,下丘脑刺激脑垂体释放FSH和LH,此时FSH浓度最大。FSH和LH可刺激卵巢中卵泡(follicle)的生长并由卵泡产生雌激素。当血液中的雌激素含量达到峰值时(标志着卵泡的成熟)
雌激素就会对脑垂体发生负反馈效应并减少FSH的释放。但雌激素对LH却会产生正反馈效应,随着雌激素浓度的增加,脑垂体会释放更多的LH,从而使LH成为脑垂体分泌最多的激素。在自发排卵的哺乳动物中,卵泡成熟后,卵大约是在LH浓度达到高峰时被释放。成熟的卵泡在LH和促乳素(prolactin)的持续影响下分泌雌激素和孕酮( progesterone ) ,但孕酮在血液中浓度的增加会对脑垂体产生负反馈效应,而后者则会导致LH释放逐渐减少。
雌激素和孕酮也会引起动物行为发生变化。各种动物在其生殖生理和内分泌学方面是各不相同的,但对哺乳动物来说,雌性个体通常是在动情周期时才接受雄性个体的求偶和交配,这一过程主要是受两种卵巢类固醇激素的影响。动物复杂的生理和行为变化是几种内分泌通道交互作用的产物,均与正反馈和负反馈系统有关。
四、激素的功能之一——激活效应(P130)
激素对行为的影响大体可区分为两类,一类是激活效应( activational effects),一类是组织效应(organizational effects)。在前者中,激素对行为的表达和表现来说好像是触发器,具有启动作用。当激素的分泌能引起一个快速反应时就是种直接的激活效应,而间接的激活效应则需要更复杂的刺激和激素分泌程序。激素的组织效应是在生物发育期间表现出来的,例如:动物的性分化和身体组织的生长格局都受着激素的调控。下面我们先就激素对行为的激活效应及其相关过程举一些实例,然后再举例说明激素的组织效应。
(略去)
5.激素与第二性征(P132)
激素除了对行为有直接的激活效应外,还可以影响动物的第二性征,公鸡受阉割后其作为第二性征的特有鸡冠就会明显减小。雄猫射尿通常是一种领地的标记行为,一旦精巢被摘除射尿行为也就随之终止。在这两个实例中,第二性征(公鸡的鸡冠)和与性别相关的行为(射尿)所发生的变化都对动物的通讯行为有影响。
6.激素与攻击行为和性行为
当环鸽受到阉割时,它的攻击行为、求偶行为和交配行为就会大大减弱。如果往阉割环鸽下丘脑的特定部位 置入晶态睾酮丙酸盐,那上述的那些行为就会恢复正常水平,这些实验显然证实了睾酮对性行为和攻击行为的激活效应,并说明特定的脑区可以被影响性行为的睾酮所激活。
其他研究结果也表明:睾酮的存在与否可影响鸟类和哺乳动物的攻击行为,如环鸽、小鼠、大鼠、公鸡和猫等。未阉割的鸟和哺乳动物更富有攻击性,诱发战斗行为的潜伏期也比较短,战斗更加频繁。阉割术常用于各种家畜如猪、牛等,为的是减少其攻击性,便于人的驾驭。金仓鼠( Mesocricetus auratus )身体两侧有成对的胁腺,它可分泌脂肪标记环境中的物体。胁腺的发育与雄激素有关,因此测量胁腺的大小及色素沉着可间接估测雄激素在血液中的相对浓度。把4只体重相同未阉割的雄性仓鼠自断奶后便隔离饲养,当在一个大空间内把它们放到一起让其自由交往时,研究人员发现在其相遇表现和胁腺大小指数之间存在着明显的正相关性(r=0.77)。该实验的主要特点是研究人员在把金仓鼠放在一起之前先对其胁腺进行了测量,因而能够根据胁腺大小(代表雄激素浓度)对相遇表现做出预测。在一个相关实验中,研究人员往每只被阉割的雄仓鼠体内注入了睾酮丙酸盐,但每只金仓鼠所注入的剂量不同。当让一组中的4只仓鼠自由交往时,研究人员可再次根据剂量的不同和相应的胁腺大小指数对相遇表现做出预测(r=0.81)。有趣的是,研究人员也可以根据雌性金仓鼠胁腺的测量结果对雌鼠相遇的行为表现做出预测。
五、激素的功能之二——组织效应
对鹌鹁、斑马雀(taeniopygia guttata)、荷兰猪、鼠和猕猴及其他动物所进行的研究清楚地表明:某些激素对动物早期发育时的性别分化有着重要影响。对哺乳动物的研究主要集中在生殖激素的组织效应,因为这些激素影响着以后成年个体的性行为和攻击行为。对鸟类的研究则主要涉及生殖激素对以后性行为的组织效应。下面我们举出一些研究实例以便说明激素对动物行为及其相关过程所施加的组织效应。
1.激素对性行为和攻击行为的影响
如果雄鼠在出生后的4 ~5天内被阉割,那么在它发育到成年期时就不再有正常的性行为。如果当这只被阉割的雄鼠发育到成年时往其体内注入雌激素和孕激素,那么它的行为表现就和雌鼠无异,例如它会做出脊柱前凸的动作,这是雌鼠接受雄鼠爬跨和插入的典型动作。如果雄鼠发育成熟后再阉割,那么注入雌激素和孕激素就不能引发它表现出雌鼠的性行为。在一次实验中往新生的雄性鼠崽中注入雌激素,当它成年后将其宰杀,经组织检查发现其精子的精巢小管发生了退化。虽然这些雄鼠仍表现有爬跨行为,但其行为很不正常,爬跨常常失准,而且不能射精。在出生后的4 ~5天内用人造雄激素或人造雌激素处理过的雌鼠,长大后不能表现出正常的月经周期。即使往其体内注入雌激素和孕激素对雄鼠的爬跨行为也毫无反应。当这些初生时就用性激素处理过的雌鼠在发育成熟时再注射人工雄激素,它就会表现出类似雄鼠的性行为。初生时用雌激素处理过的雌鼠长大后月经周期会变得不规律,若用人工雄激素处理则会发生永久性的阴道角质化。被雄性化了的雌鼠攻击倾向增加,战斗潜伏期缩短。
关于激素对雌性个体行为组织效应的类似研究还曾用豚鼠和猕猴做过,在妊娠期间用雄激素处理过的个体,其雌性后代具有雄性化的外生殖器(阴道口较小,阴蒂肥大),其性行为也类似雄性个体。
这些资料表明:对于发育早期的激素注射来说存在一个关键时期,就鼠类来说这个关键期是在出生后的4或5天内,但对豚鼠和猕猴来说这个关键期是在出生前,对新出生的豚鼠和餐进行激素注射,其效果与鼠类是不一样的。猕猴和豚鼠出生前的发育状态约相当于鼠类出生后的发育状态,所以在这些不同种类的动物中,注射激素的组织效应相对说来都发生在大体相同的发育阶段上。至于激素对鹌鹁和斑马雀生理和行为的组织效应也存在着关键期,就鹌鹁来说,雌激素组织效应的关键期是在孵化后的前两周内。
组织效应的一个有趣方面是在近年来才发现的,即并非是生理和行为的所有方面都受到同样的影响。例如不同组的鹌鹁在胚胎发育的第9天或第14天用三种不同剂量(0,5或25 μg)的雌二醇 (estradiol) 进行处理,待孵化后的第4天进行阉割以便排除孵化后释放激素的混淆效应,当发育到成年时再给予外源性睾酮。结果在第9天给予处理的雄鸟(而不是第14天)表现出了去雄性化 (demasculinized) 的性行为,而且泄殖腔腺增长加快。但无论是第9天还是第14天处理的雄鸟,其啼鸣活动都有所减弱且血液中的LH浓度降低。可见,去雄性化过程并不完全局限于一-个特定的关键期,不同的行为可能是在发育的不同时间受到影响的。
最近对3个月的爬行动物所做的研究表明:在花斑壁虎( Eublepharis macularius)、密河鳄(Alligator mississippiensis )和鳖( Trionyx spiniferus)中,行为的组织都是靠激素的组织效应实现的。如果在卵孵化前的适当年龄往这些爬行动物的卵中注入雌二醇,那么在雌激素的作用下,所有被处理的卵都会发育为雌性个体。可见,雌二醇显然会导致各种爬行动物生殖器官的雌性化。
2.激素对其他行为和胎位的影响
激素除了对动物的性行为和攻击行为有组织效应外,对其他行为也有组织效应,Meany及其同事(1982)曾研究过糖皮质激素( gucocrioids)对挪威鼠幼鼠玩耍打斗行为的影响,这种激素可降低雄性幼鼠之间玩耍-打斗的激烈程度,使其不像成年雄鼠那样具有伤害性。但对雌性幼鼠作同样的处理则不会对它们的玩要打斗行为产生影响。可见糖皮质激素对鼠类玩耍-打斗行为的组织效应是因性别而异的。
家畜(如牛)龙凤胎中的雌性个体,成长之后往往不能生有后代,据认为是与激素的作用有关,即在胚胎发育的早期,两性胎儿的血液循环是互相连通的,因此雄性激素会进入雌性胎儿体内,引起成年后的不育。最近有很多人研究了出生后用雄激素处理孪生雌仔鼠对其行为的影响,方法是在孪生雌仔鼠3月龄时用丙酸睾酮处理100天,在停止处理后的3~12个月内对其行为进行测定,结果发现:处理鼠与对照鼠相比更具有竞争优势,但两者在外貌和正常的性周期方面并无差异。这表明:早期的激素处理对动物的行为有着持久性影响,处理鼠在与其他陌生鼠相遇时比较胆大和不害怕。类似的雄激素处理实验在赤鹿中也获得了相似的结果。

在大鼠和小鼠中,雌性胎儿的胎位对其生殖结构和行为也有影响,这些鼠在每个生殖周期要排放很多卵并单育1-20个幼息,它们都有双角子宫,胎儿在子宫中是按顺序排列的,因此每一个胎儿都可能位干两个其他同性胎儿之间,或位于两个异性胎儿中间,或位于一雄一雌胎儿之间。大鼠和小鼠在妊娠后期分娩前后有一个生产和释放雄激素的短暂期,子宫中的雌鼠崽可能受到来自相邻鼠崽分泌睾酮的影响而雄性化。据研究,肛门与生殖孔之间的距离(肛-殖距离)是受雄激素影响大小的一个很方便的衡量尺度,那些在子宫中位于两雄鼠之间的雌鼠,其肛-殖距离要比位于两雌鼠之间的雌鼠大。此外受影响的还包括一些行为和生理特征。在大鼠中因在子宫中的胎位而遭到雄性化的雌鼠表现出较强的爬跨行为,面在小鼠中则表现为月经周期延长、攻击行为更强、较早衰老、日活动水平下降和更多地表现有类似雄性个体的性行为。有趣的是,胎位位于两雌胎儿之间的雄鼠具有较多的雌二醇(estradiol) ,活动水平较高,性行为较弱,而且生殖结构和脑结构也有改变。
从上述事实和实验不难看出,生殖激素对动物行为的组织的确具有影响。在早期发育的适当时刻产生的性激素(包括雄激素和雌激素)借助于产生类似雄性的个体或类似雄性的行为而影响着正在分化的脑,这一切与实际的遗传性别无关。早期性激素的分泌显然会使脑对在血液中循环的其他激素变得更加敏感,并使动物产生特定的生理和行为反应。
六、激素、环境与行为之间的相互作用(P135)
激家的激活效应常常涉及行为,激素和特定环境刺激三者之间的复杂相互作用,下面我们以实例来详细说明这种复杂的互动关系。
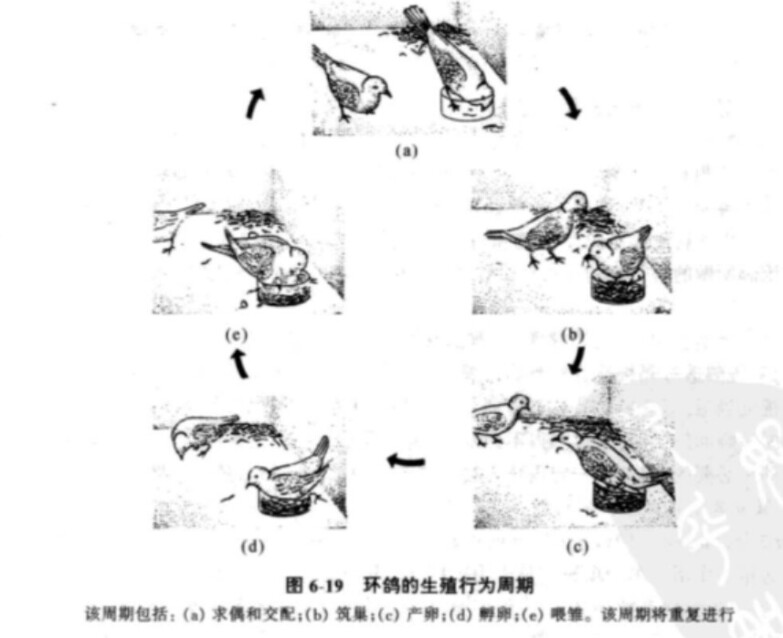
1. 环鸽生殖行为的激素调控
环鸽的整个生殖程序如图6-19所示。把雄鸽与雌鸽放在一起不久,雄鸽便开始向雌鸽求偶。被阉割的雄鸽没有求偶表现,这说明雄激素对这一生殖程序的开始是必不可少的。 雄鸽的求偶行为可刺激雌鸽的脑下垂体释放促卵泡激素(FSH),而FSH可促使卵巢中的滤泡发育,滤泡可分泌雌激素使环鸽在1 ~2天内便可开始筑巢。巢的存在本身可刺激雌鸽和雄鸽生产和分泌黄体酮。黄体酮的功能之一就是促使雌雄鸽的孵卵行为,产卵则是由雌鸽脑下垂体分泌的促黄体激素(LH)所激活的。在雄鸽体内黄体酮和睾丸素的作用刚好相反,它可抑制求偶行为和侵犯行为,而代之以孵卵行为。
一、 斑马雀(taeniopygia guttata)鸣叫行为的控制机制(P144)
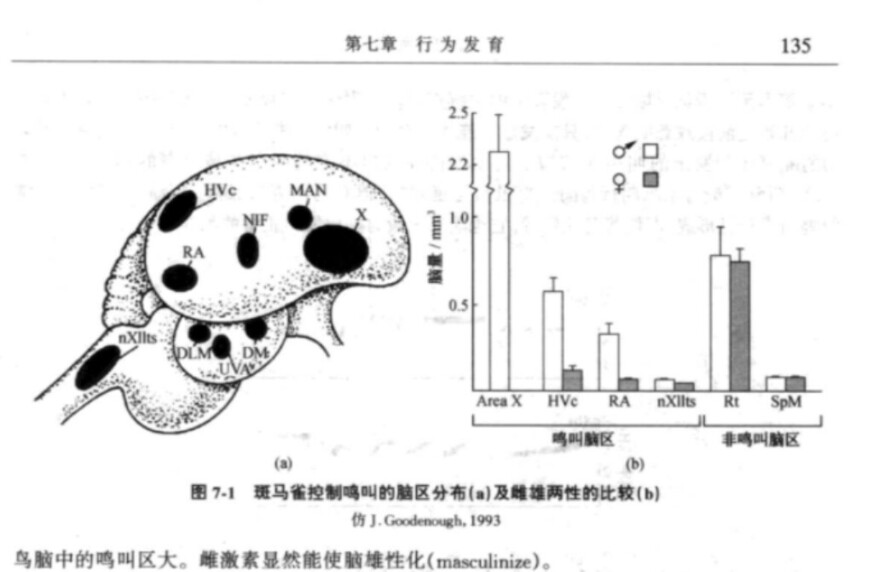
与大多数鸣禽一样,斑马雀只有雄雀能叫,其叫声是为了刺激雌鸟的生殖行为。虽然大多数种类的雄鸟可用叫声标记自己的领域,但斑马雀是群居的,可允许其他个体近距离接近自己的巢而不发生冲突。斑马雀鸣叫行为的性别差异反映着神经解剖学的两性差异,目前已确知相当大的一部分脑与控制鸣叫发声有关。鸣声通路是由9个脑区组成的,它们大都位于前脑,有条通路是支配鸣管(syrinx)发声的神经通路。 雄斑马雀的鸣叫控制区明显大于雌雀。在细胞水平上两性神经解剖学的差异也很明显,在雄雀大脑的鸣叫区,神经元比雌雀多而且大。
然而这种差异是怎样产生的呢?很显然,斑马雀两性脑结构的巨大差异是由于两性染色体的差异。雄鸟有2个X染色体[1],而雌鸟则有1个X染色体和1个Y染色体。Y染色体的缺失将导致胚胎期的雄性生殖腺生产雌激素(estrogen),这种激素可刺激大脑鸣叫区的生长与分化。实验表明,雌激素对控制鸣叫行为的神经网络有很大影响。如果把含有雌激素的药丸植入新孵化的雄斑马雀体内,它们会毫无影响(这就是说在成年期用雌激素处理的雄雀,其脑和叫声都与未处理的雄雀相似)。相反,用雌激素处理的雌雏可使脑的鸣叫区扩大,但仍不如雄鸟脑中的鸣叫区大。雌激素显然能使脑雄性化(masculinize)。[2]
其他实验也表明:激素对斑马雀的脑和鸣叫行为的影响并不限于孵化前后,虽然斑马雀的雄鸟都具有雄性化的大脑,但血液中较高的睾酮浓度是激发成鸟鸣叫的必要条件。那些孵化后不久就被植入了雌激素因而脑的鸣叫区扩大了的雌鸟,发育到成鸟时并不会鸣叫,除非给予睾酮。在孵化时未接受雌激素植入但在成年期接受睾酮的雌斑马雀也不会鸣叫。显然,雌雀在发育早期接受雌激素处理可增加成鸟脑对睾酮的敏感性并能激活其鸣叫行为。
鸟类的脑对于雌激素的组织效应并不总是敏感的,例如用雌激素处理成年雌鸟并不能影响脑内鸣叫区的大小。似乎在孵化期前后存在着一个敏感期( sensitive period),此时脑对激素的影响特别敏感。在这个时间窗内,雌激素对正在发育的神经系统影响最大。
检查文档
对文档执行检查:最后由 @ne_0x3e 在 2023-08-26T14:49:10Z 编辑